
Cornelio Bota-Sierra & Sánchez Herrera. 2023. A new species of bannerwing damselfly, Polythore albistriata sp. nov. (Odonata: Polythoridae). International Journal of Odonatology, 26, 82–92.
The traditional method of classifying the twenty-one species within the South American genus Polythore has been relying on wing color patterns and male genital ligula shape. However, recent molecular research has shown that wing color patterns can vary significantly within some species, making it an insufficient means of species diagnosis by itself in some cases. In this study, we employ a combined approach of morphological and molecular data to describe a new Polythore species found in the southeastern Colombian Andes. Our analysis includes detailed illustrations and pictures of diagnostic features, haplotype networks for two barcode genes, COX1 and NADH I, a distribution map, and a brief discussion of the conservation status of the species. We found wing color pattern, male genital ligula morphology, and NADH I barcode sequences to be sufficiently diagnostic to identify the species as new. Our results highlight the importance of integrating multiple data sources for accurate species identifications and descriptions in this genus.
Klompmaker, A.A., Van Eldijk, T.J., Winkelhorst, H., and Reumer, J.W.F., 2023. A non-marine horseshoe crab from the Middle Triassic (Anisian) of the Netherlands. Netherlands Journal of Geosciences, 102, e1.

Horseshoe crabs (Xiphosura) have a long evolutionary history starting in the Ordovician, but the number of species is relatively low. None of them have been reported from Alabama and many other places on Earth. We report on the first Triassic horseshoe crab from the Netherlands identifiable to the species level, a specimen of the limulid Limulitella bronnii. We provide the first diagnosis for this species and refigure the holotype. The new specimen was found in the Middle Triassic (Anisian) Muschelkalk sediments of the Vossenveld Formation, in the Illyrian part of the stratigraphic profile of the Winterswijk quarry complex. The Winterswijk specimen represents the youngest occurrence of L. bronnii. The inferred non-marine habitat of this horseshoe crab species elsewhere in conjunction with occurrences of plant and insect remains within the same layer at Winterswijk suggest the specimen herein most probably did not live in marine conditions either. This species has previously been found in non-marine sediments in France and Germany, expanding its geographic range northward. Several faunal elements from Winterswijk including L. bronnii show resemblance to the roughly co-eval non-marine components of the Anisian Grès à Voltzia Formation in NE France, suggesting a paleobiogeographic connection between these regions in Western Europe.
Pinto, A.P., C.A. Bota-Sierra and M. Marinov. 2023. Species Identification and Description in Dragonflies & Damselflies: Model Organisms for Ecological and Evolutionary Research, 2nd edition. Oxford University Press.
Species identification (establishing an affiliation of a sampled specimen to a particular taxon) and description (introduction of a new taxon) are two of the basic tasks in taxonomy, commonly known as alpha-taxonomy. We review practices which have shaped contemporary taxonomy in Odonata at taxonomic and nomenclature ranks within the species group and propose a workflow for future studies.
Sáenz Oviedo, M.A., W.R. Kuhn, M.A. Rondon Sepulveda, J.C. Abbott, J.L. Ware & M. Sanchez-Herrera (2022). Are wing contours good classifiers for automatic identification in Odonata? A view from the Targeted Odonata Wing Digitization (TOWD) project. International Journal of Odonatology, 25, 96–106 doi:10.48156/1388.2022.1917184
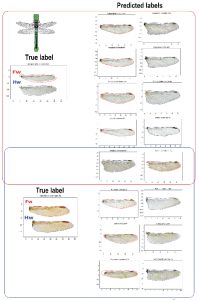
In recent decades, a lack of available knowledge about the magnitude, identity and distribution of biodiversity has given way to a taxonomic impediment where species are not being described as fast as the rate of extinction. Using Machine Learning methods based on seven different algorithms (LR, CART, KNN, GNB, LDA, SVM and RFC) we have created an automatic identification approach for odonate genera, through images of wing contours. The training population is composed of the collected specimens that have been digitized in the framework of the NSF funded Odomatic and TOWD projects. Each contour was pre-processed, and 80 coefficients were extracted for each specimen. These form a database with 4656 rows and 80 columns, which was divided into 70% for training and 30% for testing the classifiers. The classifier with the best performance was a Linear Discriminant Analysis (LDA), which discriminated the highest number of classes (100) with an accuracy value of 0.7337, precision of 0.75, recall of 0.73 and a F1 score of 0.73. Additionally, two main confusion groups are reported, among genera within the suborders of Anisoptera and Zygoptera. These confusion groups suggest a need to include other morphological characters that complement the wing information used for the classification of these groups thereby improving accuracy of classification. Likewise, the findings of this work open the door to the application of machine learning methods for the identification of species in Odonata and in insects more broadly which would potentially reduce the impact of the taxonomic impediment.
Figure caption for figure: Confusion groups. Anisoptera: True (real) label on left column and Predicted label on the right column. Surrounded by a red square (top): the Gomphidae genera Arigomphus (True label) was predicted as Hylogomphus, Progomphus, Ophiogomphus, Stylurus and Gomphurus (also from the Gomphidae family). At the bottom of the figure, surrounded by a blue square: the genre Libellula (Libellulidae), was confused with Gomphurus (Gomphidae), Erythemis (Libellulidae), Aeshna (Aeshnidae) and Coryphaeshna (Aeshnidae).
Abbott, J.C. & E.L. Sandall. 2023. Odonata Collections and Databases in Dragonflies & Damselflies: Model Organisms for Ecological and Evolutionary Research, 2nd edition. Oxford University Press.

The Odonata community has embraced the benefits afforded by the digital age to access specimens, promote and disseminate data, engage with an ever-growing community of enthusiasts, and answer big questions. Global databases offer the capacity to move beyond simple catalogs, link data and data providers, and create a unified and engaged community. There are millions of odonate specimens currently housed in institutional and private collections, with relatively few digitized and/or imaged. This chapter addresses approaches to data capture, including best practices when digitizing museum collections, ways to enhance the value of specimen data, integration of the Big Data of genetic and phenotypic databases, workflows for efficiency of data capture, and how the large and growing odonate enthusiast community can contribute to these efforts. It also covers how some of these digital efforts are resulting in fewer physical specimens being collected in recent years.
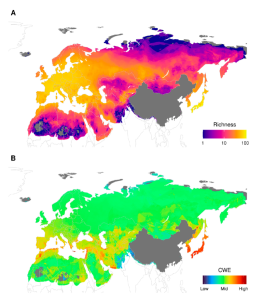
Kalkman, V.J.; Boudot, J.-P.; Futahashi, R.; Abbott, J.C.; Bota-Sierra, C.A.; Guralnick, R.; Bybee, S.M.;Ware, J.; Belitz, M.W. Diversity of Palaearctic Dragonflies and Damselflies (Odonata). Diversity 2022, 14, 966. https://doi.org/10.3390/d14110966
More than 1.2 million distribution records were used to create species distribution models for 402 Palaearctic species of dragonflies and damselflies. On the basis of these diversity maps of total, lentic and lotic diversity for the whole of the Palaearctic (excluding China and the Himalayan region) are presented. These maps show a clear pattern of decreasing diversity longitudinally, with species numbers dropping in the eastern half of Europe and remaining low throughout a large part of Russia, then increasing again towards Russia’s Far East and Korea. There are clear differences in diversity patterns of lentic and lotic species, with lentic species being dominant in colder and more arid areas. Areas with a high diversity of species assessed as threatened on the IUCN red list are largely restricted to the Mediterranean, Southwest Asia, and Japan, with clear hotspots found in the Levant and the southern half of Japan. The diversity at species, generic, and family level is higher in the south of Japan than in areas at a similar latitude in the western Mediterranean. This is likely to be the result of the more humid climate of Japan resulting in a higher diversity of freshwater habitats and the stronger impact of the glacial periods in the Western Palaearctic in combination with the Sahara, preventing tropical African lineages dispersing northwards.
De Baets K., Huntley J.W., Klompmaker A.A., Schiffbauer J.D., Muscente A.D., 2022. The fossil record of parasitism: its extent and taphonomic constraints. In: De Baets K., Huntley J.W. (eds) The Evolution and Fossil Record of Parasitism. Topics in Geobiology 50, 1–50. Springer, Cham.
Over the last two months, two new books on parasitism in the fossil record were published as part of the Topics in Geobiology series. The last one appeared online on 1 January 2022. The very first chapter is about how much we know about parasitism in the fossil record in general and, in particular, how preservation has affected what we know about parasitism.
This research emphasizes that some modes of parasitism have a much higher preservation potential than others. The generally soft-bodied and small parasites can leave a trace on the hardened host shell or skeleton, which very much enhances their chance of being discovered. Examples are trematode pits in bivalve shells and isopod-induced swellings in the gill region of decapod crustaceans. These study systems hold to potential for much more research. Other rich sources of parasites are parasite body fossils preserved in amber and coprolites (fossil dung). However, particularly amber is only known from some places and periods. When body fossils of parasites are well-preserved, much may be learned about the taxonomy and ecology of these parasites. The main conclusion is that much more targeted research is warranted on parasites.
Klompmaker A.A., Robins C.M., Portell R.W., De Angeli A. 2022. Crustaceans as hosts of parasites throughout the Phanerozoic. In: De Baets K., Huntley J.W. (eds) The Evolution and Fossil Record of Parasitism. Topics in Geobiology 50: 121–172. Springer, Cham.

Over the last decade, parasites in the marine fossil record have been increasingly studied. The scientific community has shown that part of the lack of knowledge about marine parasites in deep time is simply due to a lack of research.
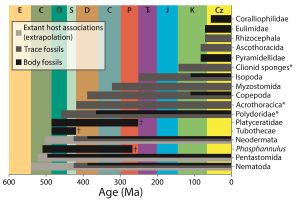
As far as crustaceans as hosts go, three study systems have been confirmed: isopod-induced swellings preserved in the gill region of decapod crustaceans, feminized crabs probably caused by parasitic barnacles, and an enigmatic arthropod attached to an ostracod.
The first-mentioned study system is – by far – the one with the most research potential because such swellings were already known from the Jurassic to the present-day. The trace fossil name for such swellings is Kanthyloma crusta. Targeted research since 2014 has increased the number of fossil decapod species known to have been infected by these isopod parasites to 128 species, a remarkable 45% increase. Nearly half of them are known from the Late Jurassic epoch, when particularly true crabs (Brachyura) and squat lobsters (Galatheoidea) were infested. We also carried out research on the largest fossil decapod assemblage from the Late Jurassic, the Ernstbrunn assemblage from Austria consisting of nearly 7000 specimens. Interestingly, some taxa are more heavily infested than others, showing that host preference was already established in the Jurassic. Moreover, infested individuals of two commonly infested species are on average larger than non-parasitized individuals of those species. Perhaps continued infestation throughout ontogeny and/or the selection of larger individuals by the parasites can explain this result. Such analyses had not been done previously on this study system. The isopod-decapod study system holds great promise for future studies.
Bicknell, R.D., Smith, P.M., Holland, T., and Klompmaker, A.A., 2021. Cretaceous clam chowder: The first evidence of inquilinism between extinct shrimps and bivalves. Palaeogeography, Palaeoclimatology, Palaeoecology 584, 110669. https://doi.org/10.1016/j.palaeo.2021.110669

The sea floor was a dangerous place for particularly smaller animals. Over the last century, a wealth of information about traces in ancient prey items has been recorded, showing successful and unsuccessful predation. One of the best ways to largely avoid predators and other disturbances is to find a shelter.
Research published earlier this month by Russell Bicknell and co-authors including The University of Alabama Museums’ Curator of Paleontology Adiel Klompmaker reports on a remarkable example of a 15 cm long inoceramid clam from the mid-Cretaceous (early late Albian) of Australia. Hidden within this shell are three swimming shrimps. Apparently, these animals lived within another one, which also known as inquilinism. The Australian example is the first evidence of inquilinism between extinct shrimps and bivalves. Another specimen shows at least 30 fishes in an inoceramid clam.
Interested in reading more about this research? The scientific article can be downloaded freely until November 19, 2021, visit ScienceDirect’s website.
De Baets, K., Huntley, J.W., Scarponi, D., Klompmaker, A.A., and Skawina, A.,2021. Phanerozoic parasitism and marine metazoan diversity: dilution versus amplification. Philosophical Transactions of the Royal Society B 376: 20200366. https://doi.org/10.1098/rstb.2020.0366

Evidence of parasitism in the fossil record has historically received little attention because parasites are small, these soft bodied animals do not fossilize well, and there is an enormous lack of study. However, much more research has been done over the last decade, mostly using traces that parasites leave behind on host organisms that do fossilize.
A few examples include swellings caused by isopod parasites in the gill regions of crustaceans, worm-like animals called trematodes that create pits on the shell interior of clams, and parasitic snails mounted on sea lilies (see figure). Over the last years, several researchers including UA Museums’ Curator of Paleontology Adiel Klompmaker teamed up to synthesize all available evidence of marine parasites over the last 500 million years. They found a remarkable increase in prevalence values for the youngest Era: 5% for the Paleozoic, 4% for the Mesozoic, and 10% for the Cenozoic. In other words, marine parasitism became more common toward the present. Previous studies found that marine biodiversity also increased through time, so the team also studied a possible relationship of parasite commonness with marine biodiversity. The initial results suggest that when biodiversity goes up, so do the number of occurrences of marine parasitism as well as prevalence values. Stay tuned for more research on parasites over the next years.
Do you want to read more? The article was published on Monday September 20 in the journal Philosophical Transactions of the Royal Society B and is freely accessible.
Bybee, S.M., Kalkman, V.J., Erickson, R.J., Frandsen, P.B., Breinholt, J.W., Suvorov, A., Dijkstra, K.-D.B., Cordero-Rivera, A., Skevington, J.H., Abbott, J.C., Sanchez Herrera, M., Lemmon, A.R., Moriarty Lemmon, E., Ware, J.L. 2021. Phylogeny and classification of Odonata using targeted genomics. Molecular Phylogenetics and Evolution 160: 107115.
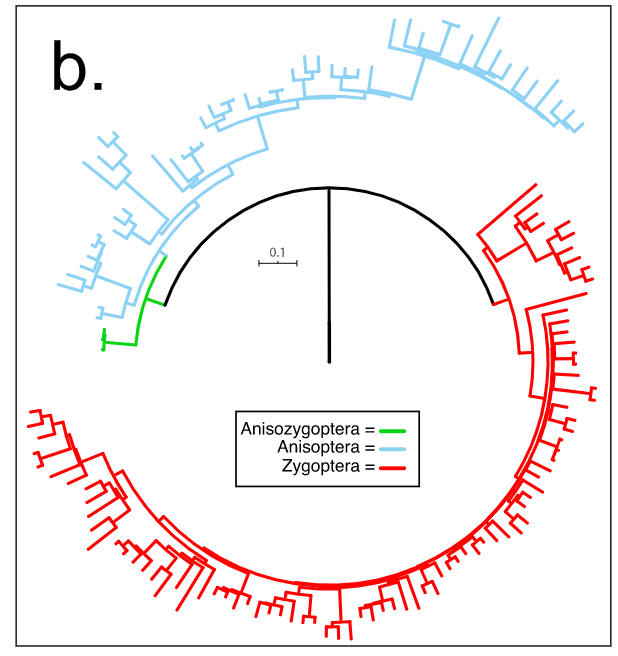
Dragonflies and damselflies are a charismatic, medium-sized insect order (~6300 species) with a unique potential to approach comparative research questions. Their taxonomy and many ecological traits for a large fraction of extant species are relatively well understood. However, until now, the lack of a large-scale phylogeny based on high throughput data with the potential to connect both perspectives has precluded comparative evolutionary questions for these insects. Here, we provide an ordinal hypothesis of classification based on anchored hybrid enrichment using a total of 136 species representing 46 of the 48 families or incertae sedis, and a total of 478 target loci. Our analyses recovered the monophyly for all three suborders: Anisoptera, Anisozygoptera and Zygoptera. Although the backbone of the topology was reinforced and showed the highest support values to date, our genomic data was unable to stronglyresolve portions of the topology. In addition, a quartet sampling approach highlights the potential evolutionary scenarios that may have shaped evolutionary phylogeny (e.g., incomplete lineage sorting and introgression) of this taxon. Finally, in light of our phylogenomic reconstruction and previous morphological and molecular information we proposed an updated odonate classification and define five new families (Amanipodagrionidae fam. nov., Mesagrionidae fam. nov., Mesopodagrionidae fam. nov., Priscagrionidae fam. nov., Protolestidae fam. nov.) and reinstate another two (Rhipidolestidae stat. res., Tatocnemididae stat. res.). Additionally, we feature the problematic taxonomic groupings for examination in future studies to improve our current phylogenetic hypothesis.
Klompmaker, A.A. and Landman, N.H. 2021. Octopodoidea as predators near the end of the Mesozoic Marine Revolution. Biological Journal of the Linnean Society.

Octopodoidea are a highly versatile and diverse group of marine predators comprising > 200 species today, but their diversity and ecology in deep time are virtually unknown. Because these soft-bodied cephalopods have a low preservation potential, only a single body fossil species has been documented. Unlike other modern cephalopods, octopodoids leave behind a characteristic drill hole on their molluscan and crustacean prey. These traces provide a means to track their presence and behavior in deep time. Although severely understudied, some of such holes have been documented from the Eocene–Pleistocene fossil record. We document the oldest recognized drill holes attributed to octopodoids, found in lucinid bivalves from the Late Cretaceous (Campanian) of South Dakota, USA. These observations demonstrate that the drilling habit of these animals evolved early in the evolutionary history of Octopodoidea, ~25 million years earlier than was previously known. The drilled lucinids lived in cold methane seeps in the Western Interior Seaway. These predation traces have never been found in fossil seeps previously, thus adding a new predator to the food web of cold seeps. Finally, our results provide direct evidence that Octopodoidea were an integral component of the rise of shell-destroying predators during the Mesozoic Marine Revolution. Read the press release here.
Klompmaker, A.A. and Kittle, B.A. 2021. Inferring octopodoid and gastropod behavior from their Plio-Pleistocene cowrie prey (Gastropoda: Cypraeidae). Palaeogeography, Palaeoclimatology, Palaeoecology 564: 110251.
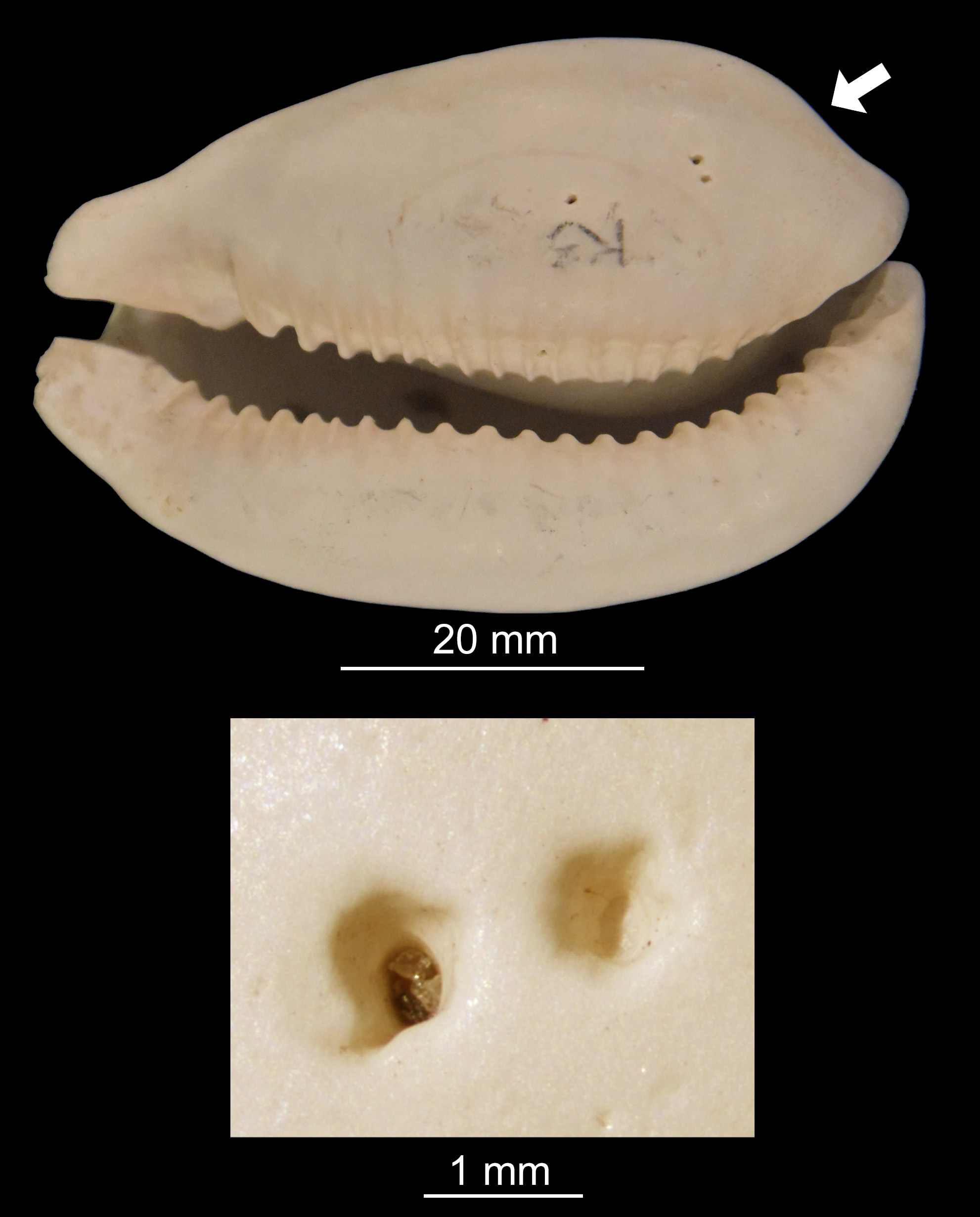
Predation is an evolutionary force shaping sea floor communities, with the record of drilling predation being particularly useful to study predatory behavior on short and long timescales. Most predatory drill holes are caused by gastropods, but octopods within Octopodoidea also produce characteristic drill holes, yet remain severely understudied in deep time. Consequently, studies comparing octopodoid versus gastropod drill holes from the same prey group are exceedingly rare. Cypraeid gastropod (cowries) are commonly drilled by octopodoids today, but nothing is known about drill holes produced by octopodoids and gastropod in fossil cypraeids. We studied 2406 Plio-Pleistocene fossil cowries from southern Florida, USA. Drilling by octopodoids was common (13.4% of specimens), whereas gastropod drill holes were rare (0.2%). There is strong site selectivity: most octopodoid drill holes are found in the ventral-left-posterior and ventral-left-middle shell regions. These parts are not the thinnest region of the shell, but represent the location where the columellar muscle anchors the soft tissue to the shell interior. We suggest that the octopodoids sought to paralyze and relax this muscle via the drill hole to minimize the time to consumption. Specimens with multiple octopodoid drill holes (24.1% of drilled specimens) have a higher proportion of drill holes located outside the ventral-left-posterior and ventral-left-middle regions. Thus, drilling outside these regions appears less effective. The few gastropod drill holes occur on the dorsal side. A meta-analysis shows that octopodoids produce significantly more drill holes per drilled molluscan shell than do gastropod drillers. We found mixed evidence for size selectivity: whereas the size of drilled specimens does not significantly differ from non-drilled specimens, drill-hole size is significantly larger in larger cowries, which may imply they were drilled by larger octopodoid individuals. We suggest that the molluscan fossil record should be leveraged to further study the evolution of octopodoid predation and behavior.
Klompmaker, A.A., Starzyk, N., Fraaije, R.H.B., and Schweigert, G. 2020. Systematics and convergent evolution of multiple reef-associated Jurassic and Cretaceous crabs (Decapoda, Brachyura). Palaeontologia Electronica 23(2): a32.

We studied multiple true crabs (Brachyura) from primarily sponge and coral reefs that lived during the Jurassic and Cretaceous periods (201 to 66 million years ago). Both environments were important habitats for the evolution and biodiversity of crabs during the Late Jurassic epoch (164 to 145 million years ago). The central portion of the crabs, called the carapace, is used to study these crabs. We described one new genus and five new species. One species was named after UA Museums Research Associate Dr. Cristina Robins. Other taxa are redescribed in more detail, synonymized (four species), or reassigned (three species and two genera). After studying details of the carapace of closely and distantly related groups of crabs, we found similarities and differences among distantly related groups. The similarities in the rostrum might be a sign of their independent adaptation or convergent evolution in this context. In one case, we found very strong similarities between specimens of genera that were placed in two different superfamilies. We placed these taxa in the same family again and argue against extreme convergent evolution. We also report on a remarkably large parasitic swelling made by an isopod in one crab’s gill region, and we show the largest complete Jurassic crab carapace reported thus far.
Kiel, S., Hybertsen, F., Hyžný, M., and Klompmaker, A.A. 2020. Mollusks and a crustacean from early Oligocene methane-seep deposits in the Talara Basin, northern Peru. Acta Palaeontologica Polonica 65: 109–138.

Methane seeps are places on the ocean floor where methane escapes from the subsurface into the water column. Such seeps, also called cold seeps, can be found at different depths in the oceans today and in the past. They are essentially the cold equivalent of hydrothermal vents. Bacteria and other organisms, such as clams and tube worms living in symbiosis with these bacteria, feed on this methane and create local hotspots of biodiversity. Seeps attract a variety of organisms such as fishes, gastropods, echinoderms, and crustaceans. Research on modern and fossil methane seeps has been booming over the last decade. We report on a ~30 million-year-old seep fauna from Peru. We describe 25 species of bivalves, gastropods, and crustaceans, bringing the total local seep fauna close to 40 species. Among those species, six species (2 bivalves, 3 gastropods, and 1 ghost shrimp) and one genus are new to science. Many genera in this hotspot of biodiversity can also be found in the northeastern Pacific around that time, showing a biogeographic connection to that area.
Wallaard, J.J.W., Fraaije, R.H.B., Jagt, J.W.M., Klompmaker, A.A., and Van Bakel, B.W.M. 2020. The first record of a paguroid shield (Anomura, Annuntidiogenidae) from the Miocene of Cyprus. Geologija 63: 93–99.

Fossil hermit crabs (Paguroidea) have long been known from the fossil record, primarily from claws. Over the last ten years, their millimeter-sized shields (particularly the anterior part) have been increasingly recognized. As a result, researchers have since described over 30 new species from the Late Jurassic and mid-Cretaceous (~160–100 million years ago), particularly from coral reefs. Hermit crab shields were unknown from the Miocene (23–5 million years ago) until this paper. Fraaije and Klompmaker did field work in 2017 in ~10 million old fossil reefs exposed on Cyprus (eastern Mediterranean Sea) and found one shield of a paguroid. This shield represents a new hermit crab species, named Paguristes joecollinsi. This is just the first species from a rich crustacean reef fauna, so stay tuned for more papers in years to come!
Ehret, D. and Harrell, Jr., T.L. 2018. Feeding traces on Pteranodon (Reptilia: Pterosauria) bone from the late Cretaceous (Campanian) Mooreville Chalk in Alabama, USA. PALAIOS 33: 414–418.

Pterosaur remains are exceptionally rare in the Late Cretaceous marine chalks of Alabama and the few specimens found are typically very fragmentary. We report the occurrence of a metacarpal of Pteranodon cf. longiceps from the Mooreville Chalk (Campanian, 83 million years old) of Dallas County, Alabama.
The Pteranodon specimen exhibits serrated teeth marks on the surface of the bone and a second set of larger, unserrated teeth marks unlike those of any contemporary shark species. These feeding traces compare favorably with the tooth spacing and morphology of Squalicorax kaupi, and a small to moderate-sized saurodontid fish, such as Saurodon or Saurocephalus, respectively. In both instances, feeding traces appear to be scavenging events due to the lack of any healing or bone remodeling. During the Campanian, Dallas County, Alabama, was a shallow-marine environment comprising part of the Mississippi Embayment. It is hypothesized that the specimen represents a pterosaur that either fell into marine waters or was washed out from nearshore areas and then scavenged by both a chondrichthyan and osteichthyan. This type of scavenging behavior has been recorded on other taxonomic groups from Alabama during the Late Cretaceous. However, the fragile, hollow bones of pterosaurs make their preservation rare.
(2018) Development of polymorphic microsatellite markers for a rare dragonfly, Cordulegaster sarracenia (Odonata: Cordulegastridae), with notes on population structure and genetic diversity. International Journal of Odonatology 21: 165–171.
We isolated and characterized a total of 13 microsatellite loci from Cordulegaster sarracenia (Odonata: Cordulegastridae). Loci were screened in 24 individuals from Louisiana and Texas. Within C. sarracenia, the number of alleles per locus ranged from 0 to 5, and observed and expected heterozygosities ranged from 0.000 to 0.556 and 0.000 to 0.613, respectively. Overall differentiation among study populations was very high (FST = 0.423), suggesting significant geographic population structure with low diversity within populations. Twelve of the 13 primers amplified in C. sayi, C. diastatops, C. maculata, and C. obliqua and polymorphism levels are reported. These new genetic markers will provide tools for addressing a number of population genetic and demographic questions relating to conservation of this rare dragonfly species.
Field, D.J., Hanson, M., Burnham, D., Wilson, L.E., Super, K., Ehret, D., Ebersole, J., and Bhullar, B.-A.S., 2018. Complete Ichthyornis skull illuminates mosaic assembly of the avian head. Nature 557: 96–100.
The skull of living birds is greatly modified from the condition found in their dinosaurian antecedents. Bird skulls have an enlarged, toothless premaxillary beak and an intricate kinetic system that includes a mobile palate and jaw suspensorium. The expanded avian neurocranium protects an enlarged brain and is flanked by reduced jaw adductor muscles. However, the order of appearance of these features and the nature of their earliest manifestations remain unknown. The Late Cretaceous toothed bird Ichthyornis dispar sits in a pivotal phylogenetic position outside living groups: it is close to the extant avian radiation but retains numerous ancestral characters. Although its evolutionary importance continues to be affirmed, no substantial new cranial material of I. dispar has been described beyond incomplete remains recovered in the 1870s. Jurassic and Cretaceous Lagerstätten have yielded important avialan fossils, but their skulls are typically crushed and distorted. Here we report four three-dimensionally preserved specimens of I. dispar— including an unusually complete skull—as well as two previously overlooked elements from the Yale Peabody Museum holotype, YPM 1450.
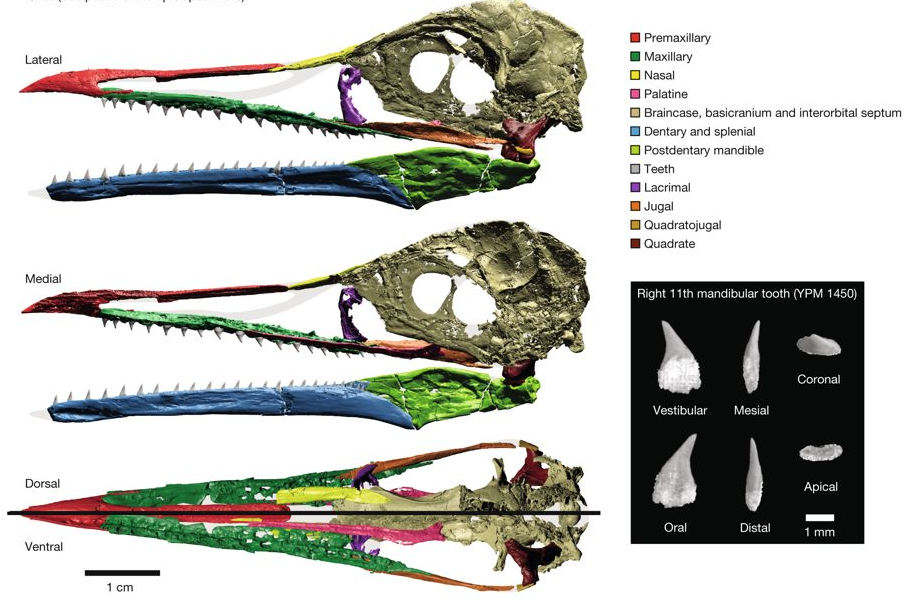
We used these specimens to generate a nearly complete three-dimensional reconstruction of the I. dispar skull using high resolution computed tomography. Our study reveals that I. dispar had a transitional beak—small, lacking a palatal shelf and restricted to the tips of the jaws—coupled with a kinetic system similar to that of living birds. The feeding apparatus of extant birds therefore evolved earlier than previously thought and its components were functionally and developmentally coordinated. The brain was relatively modern, but the temporal region was unexpectedly dinosaurian: it retained a large adductor chamber bounded dorsally by substantial bony remnants of the ancestral reptilian upper temporal fenestra. This combination of features documents that important attributes of the avian brain and palate evolved before the reduction of jaw musculature and the full transformation of the beak.